本帖最后由 rock_liberty 于 2019-5-21 02:20 编辑
0004
C-Fiber Nociceptors C纤维伤害感受器
CMHs are commonly encountered cutaneous afferents, and activity of sufficient magnitude in these fibers is thought to evoke a burning pain sensation.
C纤维-机械-热-敏感伤害感受器(CMHs)通常与皮肤的传入纤维相交汇,当这些纤维的活动强度达到足够的大小时,我们认为是其引起了灼痛的感觉。
The size of the receptive field appears to scale with the size of the animal. 感受域的大小似乎与动物的体积成比例。
Typical values for monkey are between 15 and 20 mm2 (LaMotte and Campbell 1978), and for human they are near 100 mm2 (Schmidt et al 1997). 猴子感受域的典型范围在15到20平方毫米之间(LaMotte和Campbell 1978),而对于人类,它们接近100平方毫米(Schmidt等1997)。
There are often discrete areas of mechanical sensitivity (hot spots) within the receptive field, but in many fibers the areas of mechanical responsiveness tend to fuse over the region of the receptive field. 在感受域内通常存在离散的机械敏感区域(热点),但在许多纤维的感受域中,机械响应区域倾向于融为一体。
Most CMHs respond to chemical stimuli (though not as well as A-fiber nociceptors; Davis et al 1993b) and can therefore be considered polymodal. 大多数CMH对化学刺激有反应(虽然不如A纤维伤害感受器; Davis等1993b),因此可以被认为具有多模式。
Responses to heat stimuli have been studied in considerable detail. 我们已经将对热刺激的响应进行了详尽的研究。
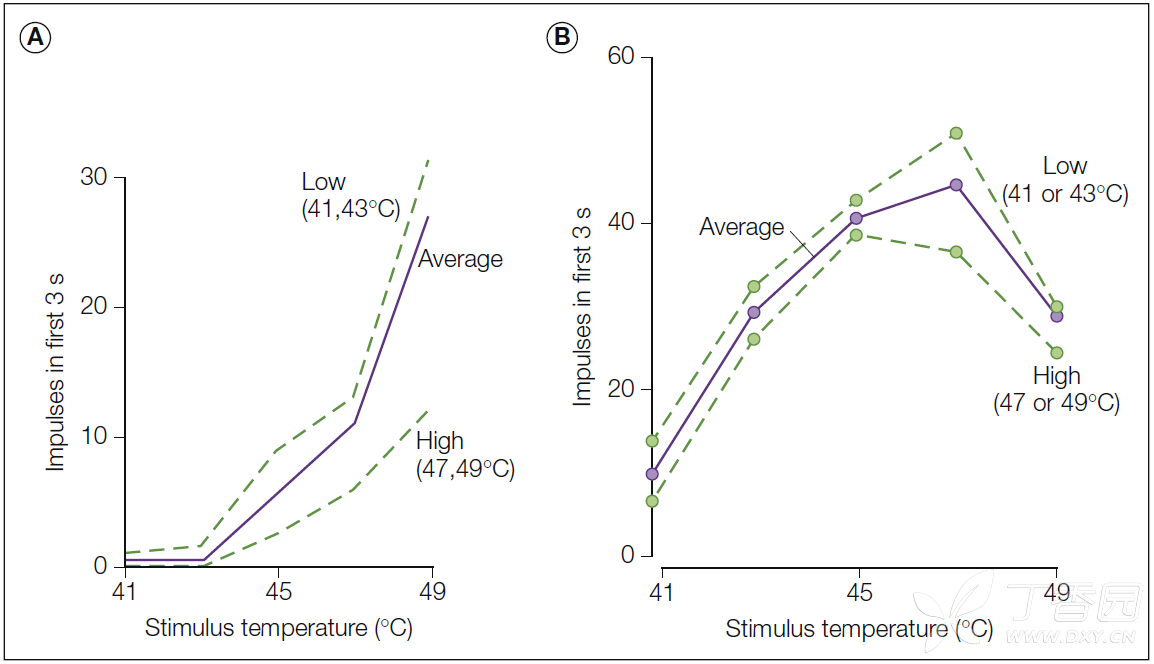
Figure 1-1. Responses of a typical C-fiber nociceptor and a warm fiber to heat stimuli. Heat stimuli ranging from 41-49°C and lasting 3 seconds were presented at 25-second interstimulus intervals to the glabrous skin of the monkey hand. Each stimulus occurred with equal frequency and was preceded by every other stimulus an equal number of times. Within these constraints, the order of stimulus presentation was randomized. Base temperature between stimuli was 38°C. A, Monotonic stimulus–response function for a typical nociceptor. B, Non-monotonic stimulus–response function for a typical warm fiber. The solid line represents the total response to a given temperature averaged across all presentations. The dotted lines represent the stimulus–response functions obtained when the preceding temperature was of low (41 and 43°C) or high (47 and 49°C) intensity. (Reproduced with permission from LaMotte RH, Campbell JN 1978 Comparison of responses in warm and nociceptive C-fiber afferents in monkey with human judgements of thermal pain. Journal of Neurophysiology 41:509–528.)
Figure 1-1. 典型的C纤维伤害感受器和温热纤维对热刺激的反应。 对猴子手部的无毛皮肤施加范围为41~49°C的热刺,持续3秒,每次刺激间隔25秒。 每一刺激的频率均一致,并且在每个其他刺激之前发生相同的次数。 在这些条件的约束下,刺激施加的顺序是随机的。 基础环境温度为38℃。 A,典型伤害感受器的单调刺激 - 反应曲线。 B,典型温暖纤维的非单调刺激 - 响应曲线。 实线表在所有给定温度的平均值下的总反应情况。 虚线表示当前一刺激的温度为低(41和43°C)或高(47和49°C)强度时获得的刺激响应曲线。(Reproduced with permission from LaMotte RH, Campbell JN 1978 Comparison of responses in warm and nociceptive C-fiber afferents in monkey with human judgements of thermal pain. Journal of Neurophysiology 41:509–528.)
The response of a typical CMH to a random sequence of heat stimuli ranging from 41–49°C is shown in Figure 1-1A. 典型的CMH对41-49°C范围内随机序列热刺激的响应如Figure 1-1A所示。
It can be seen that the response increases monotonically with stimulus intensity over this temperature range, which encompasses the pain threshold in humans. 可以看出,在该温度范围内,响应随着刺激强度单调增加,人类的疼痛阈值亦包含在其中。
One ion channel involved in the transduction of heat at nerve terminals is thought to be the neuronal transient receptor potential ion channel V1 (TRPV1); activity in this channel increases with increasing temperature (Caterina et al 1997). 一种参与神经末梢热传导的离子通道被命名为神经元瞬时受体电位离子通道V1(TRPV1); 该通道中的活性随着温度的升高而增加(Caterina等1997)。
A detailed description of the neuronal ion channels involved in stimulus transduction is presented in Chapter 2 (for review see Dubin and Papapoutian 2010). 有关刺激转导的神经元离子通道的详细描述见第2章(综述见Dubin和Papapoutian 2010)。
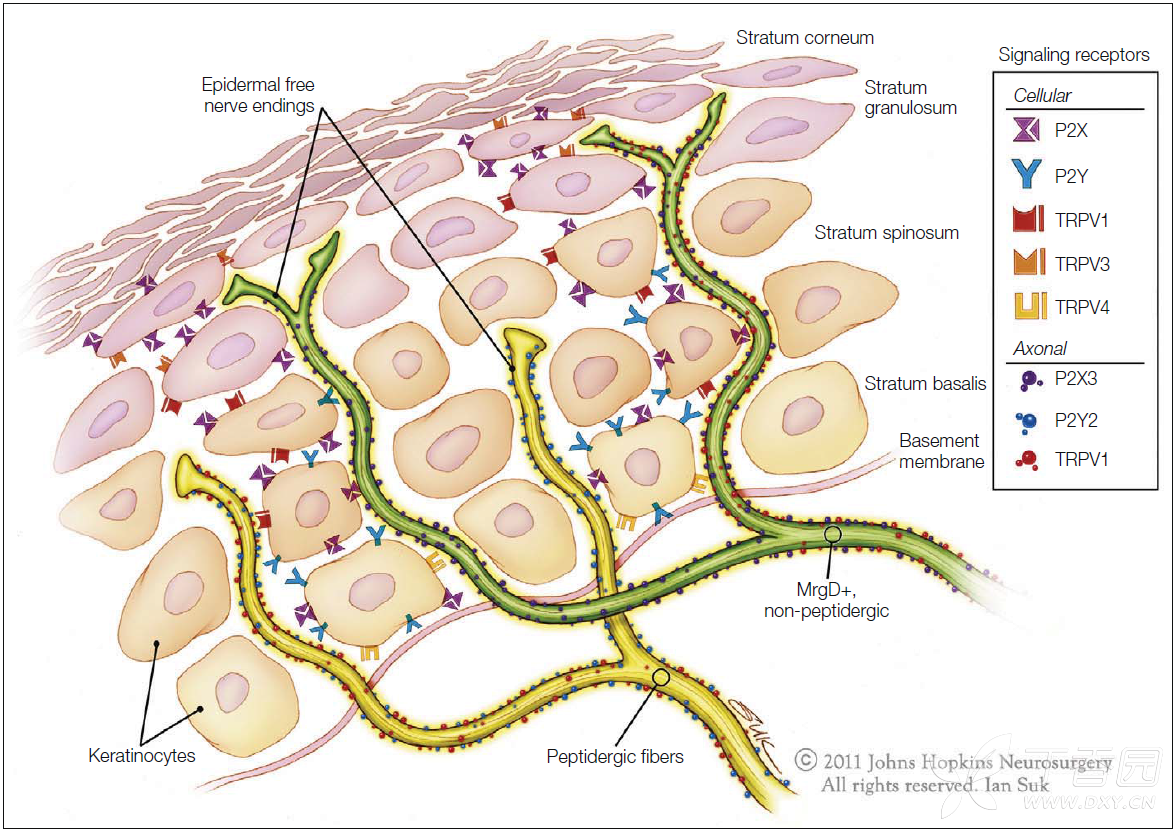
Figure 1-4. Schematic illustration of unmyelinated fiber terminations in the epidermis. Non-peptidergic, MrgD+ neurons terminate as free nerve endings in the most superficial layers of the epidermis. Peptidergic neurons terminate in deep layers of the epidermis. Some of the signaling receptors found on keratinocytes and free nerve endings are also illustrated. (Artwork by Ian Suk, Johns Hopkins University; adapted from Dussor G, Koerber HR, Oaklander AL, et al 2009 Nucleotide signaling and cutaneous mechanisms of pain transduction. Brain Research Reviews 60:24–35.)
Figure 1-4. 表皮中无髓鞘纤维末梢的示意图。非肽类,MrgD(+)的神经元以游离神经末梢的形式终止于表皮的最浅层。肽能神经元终止于表皮的深层。在角质层上发现的一些信号传导手提以及游离神经末梢亦在示意图中表示了出来。(Artwork by Ian Suk, Johns Hopkins University; adapted from Dussor G, Koerber HR, Oaklander AL, et al 2009 Nucleotide signaling and cutaneous mechanisms of pain transduction. Brain Research Reviews 60:24–35.)
Signal transduction molecules on keratinocytes may also play a role in heat transduction by inducing the release of adenosine triphosphate (ATP), which activates purinergic receptors (P2X3 and P2Y2) on the free nerve endings (see Fig. 1-4). 角质形成细胞上的信号转导分子也可通过诱导三磷酸腺苷(ATP)的释放而在热转导中起作用,其激活游离神经末梢上的嘌呤能受体(P2X3和P2Y2)(see Fig. 1-4).
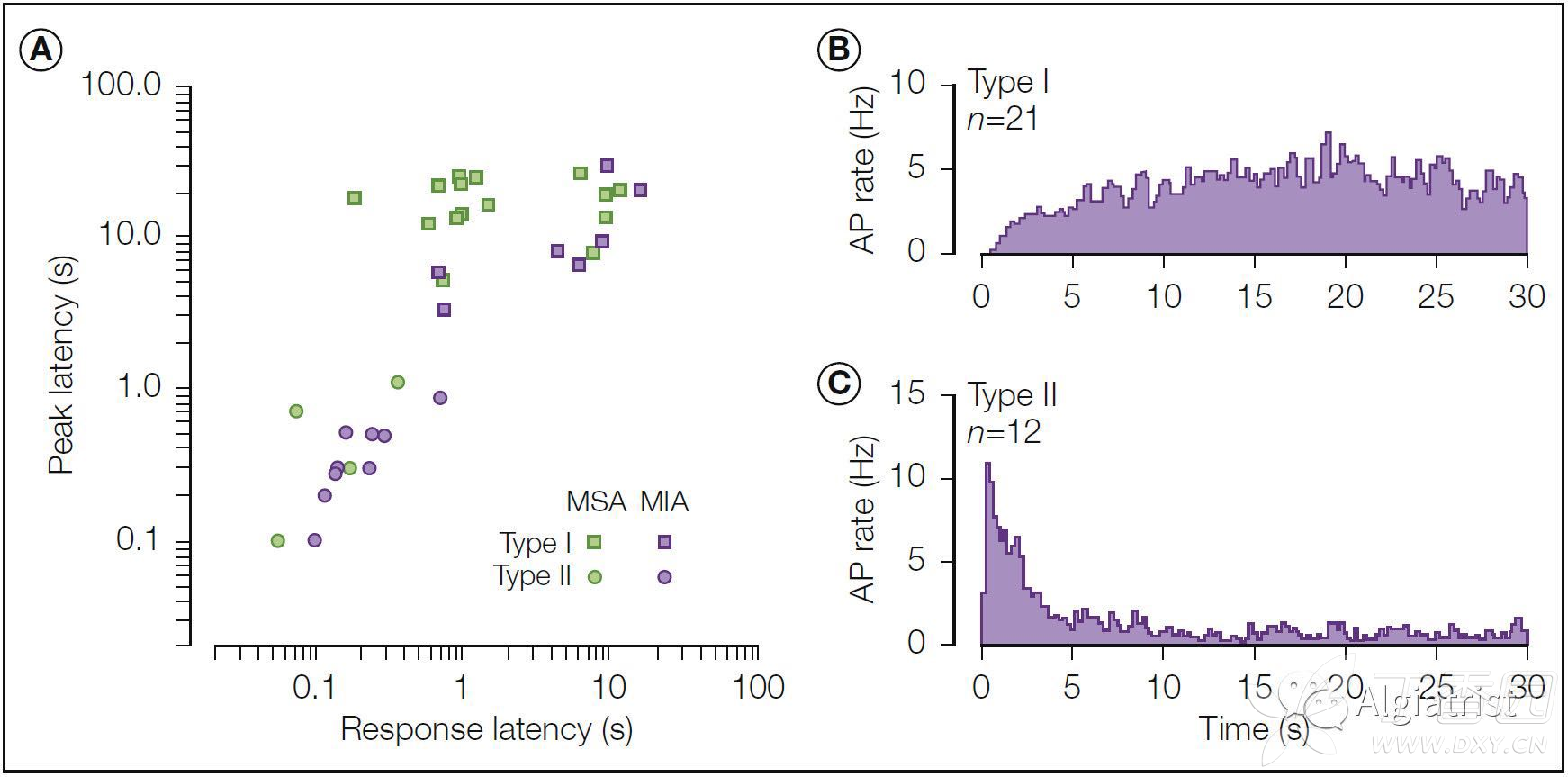
Two types of heat response are observed following a stepped heat stimulus. 在阶梯式热刺激之后观察到两种类型的热响应。
Figure 1-2. Two types of heat responses are observed in C-fiber nociceptors. A, Stepped heat stimulus (49°C, 3 seconds) used to classify heat response. B, The quick C (QC) fiber (yellow circles) exhibits a high-frequency discharge during the rising phase of the stimulus that adapts quickly (within 1 second). The slow C (SC) fiber (blue circles) exhibits a relatively uniform discharge throughout the stimulus period. Each circle represents the time of occurrence of an action potential. C, A histogram of the heat thresholds reveals that the distributions of QC and SC fibers are almost non-overlapping. (From Johanek LM, Meyer RA, Friedman RM, et al 2008 A role for polymodal C-fiber afferents in nonhistaminergic itch. Journal of Neuroscience 28:7659–7669.)
Figure 1-2.在C纤维伤害感受器中观察到两种类型的热响应。 A,阶梯式热刺激(49°C,3秒)用于对热响应分类。 B,快速C(QC)纤维是快适应型感受器(在1S内适应),在刺激的上升阶段表现出高频放电(黄色圆圈)。 缓慢的C(SC)纤维,在整个刺激期间表现出相对均匀的放电(蓝色圆圈)。 每个圆圈表示动作电位发生的时间。 C,热阈值的直方图显示QC和SC纤维的分布几乎不重叠。(From Johanek LM, Meyer RA, Friedman RM, et al 2008 A role for polymodal C-fiber afferents in nonhistaminergic itch. Journal of Neuroscience 28:7659–7669.)
Quick C (QC) fibers exhibit their peak discharge during the rising phase of the heat stimulus, whereas slow C (SC) fibers exhibit their peak discharge during the plateau phase (Fig. 1-2B).
快速C(QC)纤维的放电峰在热刺激的上升阶段出现,而缓慢的C(SC)纤维的放电峰在平台阶段表现出峰值放电(Fig. 1-2B)。
The heat thresholds (Fig. 1-2C) and mechanical thresholds of QC fibers are significantly lower than those of SC fibers, thus suggesting that they may be located more superficially in the epidermis. QC纤维的热阈值(Fig. 1-2C)和机械阈值显着低于SC纤维的阈值,因此表明它们在表皮中的位置更加表浅。
QC fibers respond more vigorously to pruritic stimuli than do SC fibers, which suggests that they may be important in itch sensations (Johanek et al 2008). QC纤维对瘙痒刺激的反应比SC纤维更强烈,这表明它们在瘙痒感觉中可能具有重要作用(Johanek et al 2008)。
Thermal modeling studies combined with electrophysiological analysis have indicated that (1) the heat threshold of CMHs depends on the temperature at the depth of the receptor and not the rate of increase in temperature, (2) transduction of heat stimuli (conversion of heat energy to action potentials) occurs at different skin depths for different CMHs (Tillman et al 1995b), and (3) suprathreshold responses of CMHs vary directly with the rate of increase in temperature (Tillman et al 1995a, 1995b; Yarnitsky et al 1992). 热调节研究结合电生理学分析表明: CMHs的热阈值取决于受体所处深度处的温度,而不是温度升高的速率 对于不同的CMH,热刺激的转换(热能转换为动作电位)发生在不同的皮肤深度(Tillman et al 1995b) CMH的超阈值响应直接随温度的升高速率而变化(Tillman et al 1995a, 1995b; Yarnitsky et al 1992)
The depth of the heat-responsive terminals of CMHs varies quite widely (ranging from 20–570 μm; Tillman et al 1995b). CMH的热响应末梢的深度变化相当广泛(范围为20-570μm; Tillman et al 1995b)。
When a stepped temperature stimulus is applied to the skin, the temperature increases in the subsurface levels more slowly because of thermal inertia. 当对皮肤施加阶梯式温度刺激时,由于热惯性,温度在深处上升得更慢。
The disparity in the surface temperature and the temperature at the level of the receptor varies directly with depth and indirectly with time. 皮肤表面温度和受体水平温度的差异直接随深度而变化,间接随时间变化。
Given that the depth of CMH terminals varies widely, true heat thresholds are obtained when the rate of increase in temperature is very gradual or when the duration of the stimulus is very long. 鉴于CMH末梢的深度变化很大,当温度增加速率非常缓慢或刺激持续时间很长时,才能获得真正的热阈值。
Although the literature reflects a wide range of heat thresholds for CMHs, when tested with these types of heat stimuli, the heat threshold of the majority of CMHs is in a remarkably narrow range of 39–41°C (Tillman et al 1995b). 尽管文献描述了CMH的各种热阈值,在用这些类型的热刺激进行测试时,大多数CMH的热阈值在39-41°C这一非常窄的范围内(Tillman等1995b)。
The response of CMHs is also strongly influenced by the stimulus history. CMH的反应也受到过去的刺激的的强烈影响。
Both fatigue and sensitization are observed. 敏化与疲劳的现象均可被观察到。
One example of fatigue is the observation that the response to the second of two identical heat stimuli is substantially less than the response to the first stimulus. 举例说明所观察到的疲劳现象,我们观察到,对于两个相同热刺激,对第二个热刺激的响应明显小于对第一个热刺激的响应。
This fatigue is dependent on the time between stimuli, with full recovery taking longer than 10 minutes. 这种疲劳的程度取决于刺激之间的时间间隔,完全恢复需要超过10分钟。
A similar reduction in the intensity of pain after repeated heat stimuli is observed in human subjects (LaMotte and Campbell 1978). 在人类受试者中能观察到,在反复热刺激后疼痛强度存在类似的降低(LaMotte和Campbell 1978)。
Fatigue is also apparent in Figure 1-1A, where the response to a given stimulus varied inversely with the intensity of the preceding stimulus.
疲劳现象在Figure 1-1A中也很明显,如果将两次刺激的响应情况进行对比,第二次刺激对应的响应的强度与第一次刺激的强度成反比。
A decrease in the response to heat is also observed following mechanical stimuli applied to the receptive field or electrical stimuli applied to the nerve trunk (Peng et al 2003). 对感受域施加机械刺激对神经干施加电刺激后,也能观察到对热刺激的响应强度降低(Peng等,2003)。
This suggests that fatigue in response to a given stimulus modality can be induced by heterologous stimulation, that is, by excitation with a stimulus of a different modality. 这就表明,通过异源刺激可以诱导出给定刺激类型所对应的响应的疲劳现象,即通过不同类型的刺激均可引发。
Interestingly, recovery from cross-modal or heterologous fatigue is faster than recovery from fatigue induced by a stimulus of the same modality. 有趣的是,相较于相同类型刺激下所引发的疲劳现象,不同刺激类型(异源刺激)引发的疲劳现象所需的恢复时间更短。
Presumably, this is because these heterologous stimuli do not activate and therefore do not fatigue the stimulus transduction apparatus in the same way. 我们推测,这是因为异源刺激是以不同的方式激活传导通路的,因而所引发的疲劳现象也有所不同。
Alternatively, fatigue may arise from independent effects on spike initiation (from antidromic stimulation) and transduction (from natural stimulation at the receptive field). 亦这种可能性:疲劳现象可能是形成动作电位(来自逆向刺激)和转导(来自感受域的自然刺激)的独立影响所引发的。
Fatigue in response to heat stimuli is also seen in vitro when small (and presumably nociceptive) dorsal root ganglion (DRG) cells are repetitively tested with heat stimuli (Greffrath et al 2002). 当在体外用热刺激重复测试小的(并且可能是伤害性的)背根神经节(DRG)细胞时,也可观察到对热刺激的响应疲劳现象(Greffrath等2002)。
The enhanced response, or sensitization, that may occur in CMHs after tissue injury is described below in the section on hyperalgesia. 组织损伤后可能在CMH中发生的敏化将在下文的痛觉过敏部分中描述。
Responses to mechanical stimuli are covered in more detail later. 对机械刺激的响应的细节将在后文中进行介绍。
Suffice it here to indicate that CMHs usually display a slowly adapting response to mechanical stimuli of a given force. 这里需要指出的是,在给定力的机械刺激下,CMH通常是慢适应的。
As noted later, MSA CMHs have a graded response to punctate stimuli, but their stimulus–response functions become saturated at levels substantially below the threshold for pain. 如后所述,MSA(机械敏感传入神经) CMH对点状刺激有分级反应,但其刺激-反应曲线在低于疼痛阈值的水平上变得饱和。
C-fiber MIAs are heterogeneous with regard to responses to chemical and heat stimuli, and some respond only to mechanical stimuli (but of course with a very high mechanical threshold). C纤维MIA(机械不敏感传入神经)在对化学和热刺激的反应方面是异质的,并且它们中的一些仅响应机械刺激(但机械阈值非常高)。
The sensitivity to mechanical stimuli has no obvious correlation to the heat threshold (Davis et al 1993b). 对机械刺激的敏感性与热阈值之间不存在明显的相关性(Davis等,1993b)。
In contrast to CMH afferents, some C-fiber MIAs in humans are vigorously excited when challenged with histamine or capsaicin. 与CMH传入相反,使用组胺或辣椒素可强烈兴奋人类的一些C纤维MIA。
In addition, the activity observed in these C-fiber MIAs parallels the duration of the perception of itch (histamine) or burning pain (capsaicin) (Schmelz et al 1997, 2000b).
此外,在这些C纤维MIA中观察到的兴奋状态与瘙痒(组胺)或灼痛(辣椒素)感知的持续时间相似(Schmelz等1997,2000b)。
C-fiber MIAs may therefore act as chemosensors. 因此,C纤维MIA可以被视为化学感受器。
In addition to pronounced chemosensitivity, these fibers have some other interesting properties that could account for pain in response to tonic pressure stimuli or the neurogenic flare response (see below). 除了显著的的化学敏感性外,这些纤维还具有一些其他有趣的特性,可以对强直性压力刺激或神经源性反应所引起的疼痛进行解释(见下文)。
Low-threshold C-fiber mechanoreceptors that do not re- spond to heat have been described in the cat (Bessou and Perl 1969) and rabbit (Shea and Perl 1985). 在猫(Bessou和Perl 1969)和兔子(Shea和Perl 1985)中,他们描述了不对热的低阈值C-纤维机械感受器。
In primates, including humans, these fibers have been found in proximal areas of the body (Kumazawa and Perl 1977, Nordin 1990) and the hairy skin on the forearm (Vallbo et al 1999). 在包括人类在内的灵长类动物中,我们已经在身体的近端区域(Kumazawa和Perl 1977,Nordin 1990)和前臂上的多毛皮肤(Vallbo等1999)发现了这些纤维。
These afferents are strongly activated by innocuous mechanical stimuli moved slowly across the receptive field, but they also respond to pinprick stimuli. 这些传入神经被在感受域上缓慢移动的无害的机械刺激强烈激活,但它们也对针刺刺激作出反应。
The neuronal activity in these fibers is not critical for the perception of touch and, according to one imaging study, leads to the activation of the insular but not the sensory cortex (Olausson et al 2003). 这些纤维引起的神经元活动对于触觉的感知并不重要,并且根据一项影像学研究,它们引起脑岛而非感觉皮层的激活(Olausson等人2003)。
Low-threshold C-fiber mechanoreceptors are thought to mediate the sensation of “pleasant” touch and may therefore play an important role in “affiliative” behavior (Vallbo et al 1999, Wessberg et al 2003, Löken et al 2009). 低阈值C-纤维机械感受器被认为介导“愉快”触觉的感觉,因此可能在“亲近”行为中起重要作用(Vallbo等1999,Wessberg等2003,Löken等2009)。
Some mechano-insensitive C fibers are reported to be activated by non-noxious and noxious cold and hot stimuli. 据报道,一些对机械不敏感的C纤维能偶被非有害和有害的冷热刺激所激活。
It has been hypothesized that activity in these afferents may mediate the “hot–burning” sensations caused by such stimuli. 已有假说认为,这些传入神经的活动可以介导由这些刺激引起的“热-燃烧”感觉。
These afferents may also be involved in mediating psychophysical phenomena such as “paradoxical heat” or the thermal grill illusion (Campero et al 2009). 这些传入神经也可能参与介导精神物理现象,如“矛盾热”或热烧烤错觉(Campero等2009)。
C-fiber afferents differ not only in their receptive features but also in their conductive properties. 不仅在作为感受器的方面的特点,C纤维传入神经在传导方面也有与众不同的特点。
In fact, their conductive and receptive properties appear to correlate. 事实上,它们的作为感受器的方面的特点和传导方面的特点似乎具有相关性。
When unmyelinated C-fiber afferents are activated repetitively by electrical stimuli, their conduction latency increases gradually (i.e., the conduction velocity of the afferent decreases). 当通过电刺激重复激活无髓鞘的C纤维传入神经时,它们的传导潜伏期逐渐增加(即传入纤维的传导速度减小)。
In addition, with increasing stimulation frequency, the amount of this activity-dependent slowing increases. 此外,随着刺激频率的增加,这种依赖于兴奋状态的减速的量同时增加。
Slowing in C-fiber MIAs is greater than in C-fiber MSAs (Weidner et al 1999), and mechanosensitive nociceptive afferents show more pronounced slowing than do cold-sensitive C fibers, low-threshold C fibers, or sympathetic efferent C fibers (Gee et al 1996, Serra et al 1999, Obreja et al 2010, Ringkamp et al 2010). C纤维MIAs比C纤维MSA的减速现象更显著(Weidner等1999),机械敏感性伤害性传入神经表现出比冷敏感C纤维,低阈值C纤维或交感神经传出C纤维更明显的减速现象(Gee等1996,Serra等1999,Obreja等2010,Ringkamp等2010)。
This difference in slowing properties indicates that the ion channels responsible for conduction may be different and suggests that the ion channels responsible for spike initiation at the receptive terminal may also differ between C-fiber classes. 这种减速现象的差异表明,负责传导的离子通道可能有所不同,并且提示我们,在感受末梢处负责形成动作电位的离子通道在不同C纤维类别之间也可能有所不同。
| 



 |关于我们|业务合作|手机版|新青年麻醉论坛
( 浙ICP备19050841号-1 )
|关于我们|业务合作|手机版|新青年麻醉论坛
( 浙ICP备19050841号-1 )





 发表于 2019-5-12 09:07:50
发表于 2019-5-12 09:07:50





 QQ好友和群
QQ好友和群 收藏
收藏 淘帖
淘帖 鲜花
鲜花 鸡蛋
鸡蛋 提升卡
提升卡 置顶卡
置顶卡 变色卡
变色卡 楼主
楼主
 发表于 2019-5-13 12:47:50
发表于 2019-5-13 12:47:50